以野韭菜为例,探讨细胞学技术在再生植株遗传稳定性的改善与应用


文字/有范知识
编辑/有范知识

体外培养的植物细胞的存在普遍的核不稳定性,这一现象在广泛的植物材料中得到了证实—— 这种不稳定性通常表现为培养物染色体以及从中再生的植物染色体发生数量和结构上的改变。
笔者将利用细胞学技术,对野韭菜再生植株遗传稳定性方面的有效性进行了实验观测,以验证细胞学技术在确认再生植物遗传稳定性方面的应用可行性和有效性。

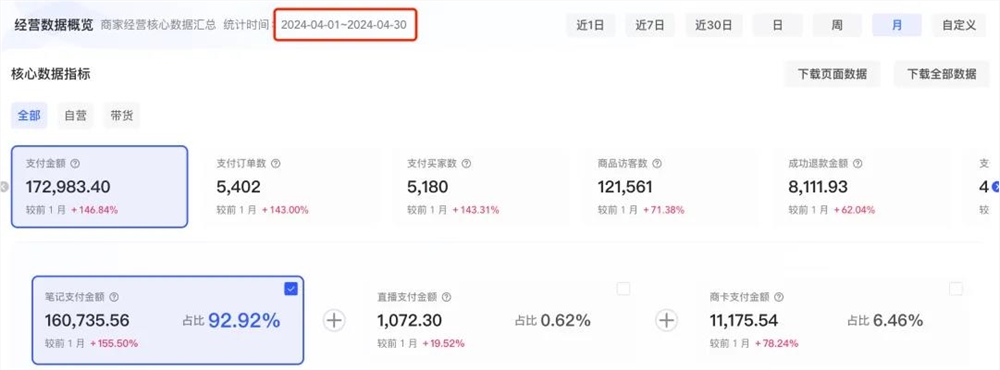
野韭菜(葱科)是花园洋葱的重要野生近缘种,野韭菜中已知存在二倍体(2n=16)和四倍体(2n=32)细胞型,是非常具有研究价值的再生植株研究对象。
近年来,由于当地居民过度开采大蒜味的花序和叶子,缺乏有组织的种植,只采不种,导致具有重要经济意义的物种的野生遗传资源在自然栖息地中一直在减少,无法实现生物资源的可持续良性发展。

基于严峻的基因灭绝威胁,国家植物组织培养库努力对这一重要物种以及其他相关类型进行体外保护。 随后成功地完成了通过体外芽增殖的克隆繁殖,尝试使用细胞学技术研究不同世代再生植物的遗传稳定性。
通过再生细胞的细胞学研究表明,大部分(>90%)的细胞具有正常的补体,变异仅在一小部分细胞中发生,如瘤变、多倍体和结构改变。
与直接再生相比,这种结构和数值畸变的频率非常高,在通过愈伤组织再生时表现得尤为突出。

不得不提的是,迁地保护为主要目标时,在直接再生和通过愈伤组织再生这两种情况下,所有克隆增殖都应经常监测和验证培养材料的基因组完整性。
除了生物化学、组织学和组织化学之外,细胞遗传学分析是确定在再生和器官发生过程中核物质是否发生任何变化的最有信息和最可靠的技术之一。
在细胞遗传学方面,核型研究最为重要,因为它们通常提供有关染色体结构、数量和总体形态的真实信息。

另一方面,减数分裂侧重于染色体配对行为的细节、重组频率的研究,更加关注染色体在后期 I阶段 和 II 阶段的分离模式,有丝分裂研究中不能直接推断出来配对细节和重组频率。
下面一起来了解野韭菜体外培育研究的试验方法、观测过程与结果分析。
关于建立体外再生小植株的标准方案已在其他文章中讨论过,在此不做赘述,下面详细介绍本次实验的材料与方法。
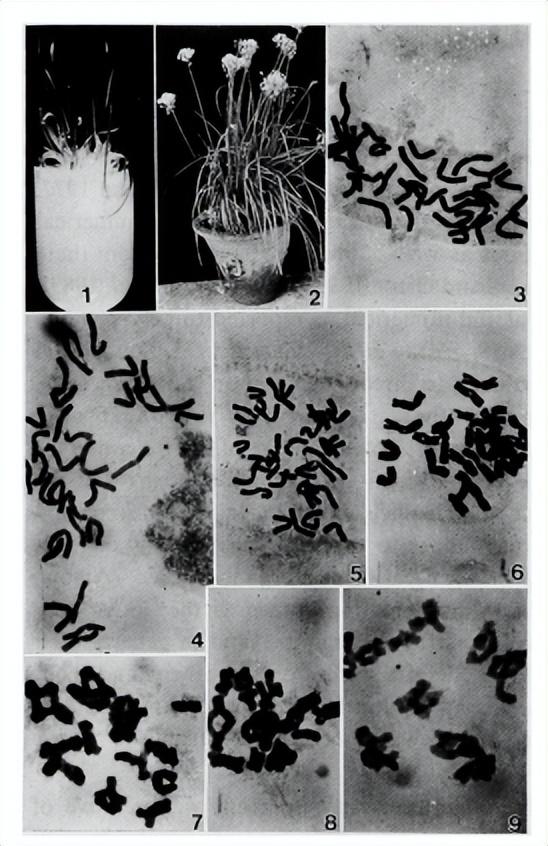
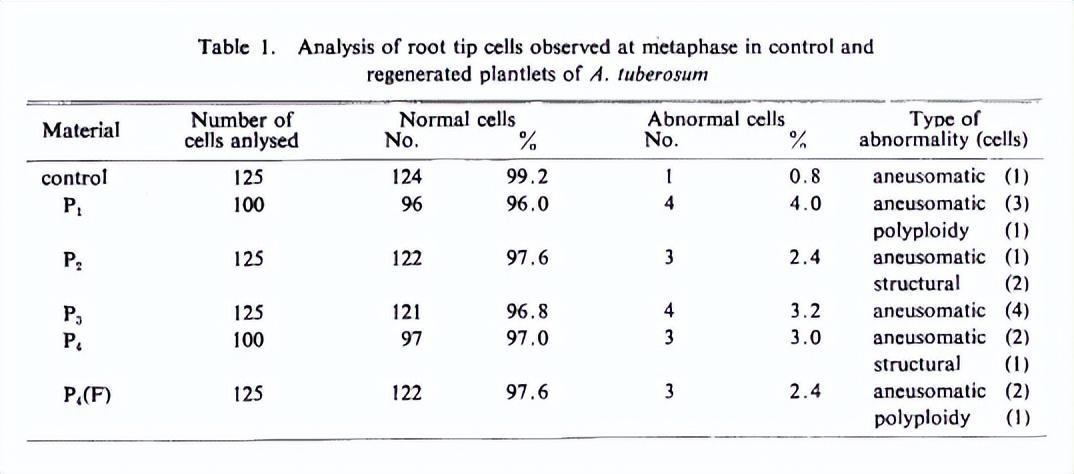
试验对体内(对照)和体外(再生植株)材料都分别进行了核素学研究,包括四个连续传代,分别标记为P1、P2、P3和P。
随后在田间建立P(母本),培养传代的植株,命名为P,(F)。为了确定核形态的变化,另外对体外培养的5个再生体进行了研究。

每个再生材料的根尖制备至少五张载玻片,平均每张载玻片上四个细胞组织,确保评分的数据偏差小于合理范围。
为了进行减数分裂分析,同时从对照植株和P4(F)植株上采集适当大小的花蕾。

采集目标样本:切除生长活跃的根尖(约1-2厘米),在4℃双蒸馏水中预处理24小时。预处理后的根尖在1:3冰醋酸:95%乙醇中固定过夜。然后将根尖保存在70乙醇中,温度为10摄氏度,直至压扁。
将花蕾直接固定在上述固定液中,并加入一滴FeCl3溶液,固定至少24小时,然后在温度10摄氏度的70%乙醇中保存。
制备染色体:在3-4次蒸馏水中洗涤材料(根尖/花药),并在60摄氏度的盐酸中水浴水解10分钟。充分洗涤后,将水解后的材料转移到浅色碱性品红溶液中,在室温下黑暗保存约45分钟。将染色材料用1%的乙酰胭脂红压碎,用ORWO MAB光负片从临时制剂中拍摄显微照片。
保存载玻片:用液氮冷冻技术去除盖玻片,并使用1:3,1:6,1:9(冰醋酸:95%乙醇)系列脱水,然后用95%乙醇进行两次变形,然后再装在D.P.X.中。对于有丝分裂和减数分裂制备的分析,仅对具有良好扁平形态和可计数染色体的细胞进行评分。
对照植物根尖细胞中分析的大多数细胞的染色体数为32,但是其中遇到了一个特殊的、仅有30条染色体的单细胞异构体(2n=30)。
在P1、P2、P3中,正常染色体数目的细胞比例极高。
在P中,96%的细胞显示正常染色体数目为2n=32,而其余4%的细胞分别在3和1细胞中被发现为非整倍体或多倍体。
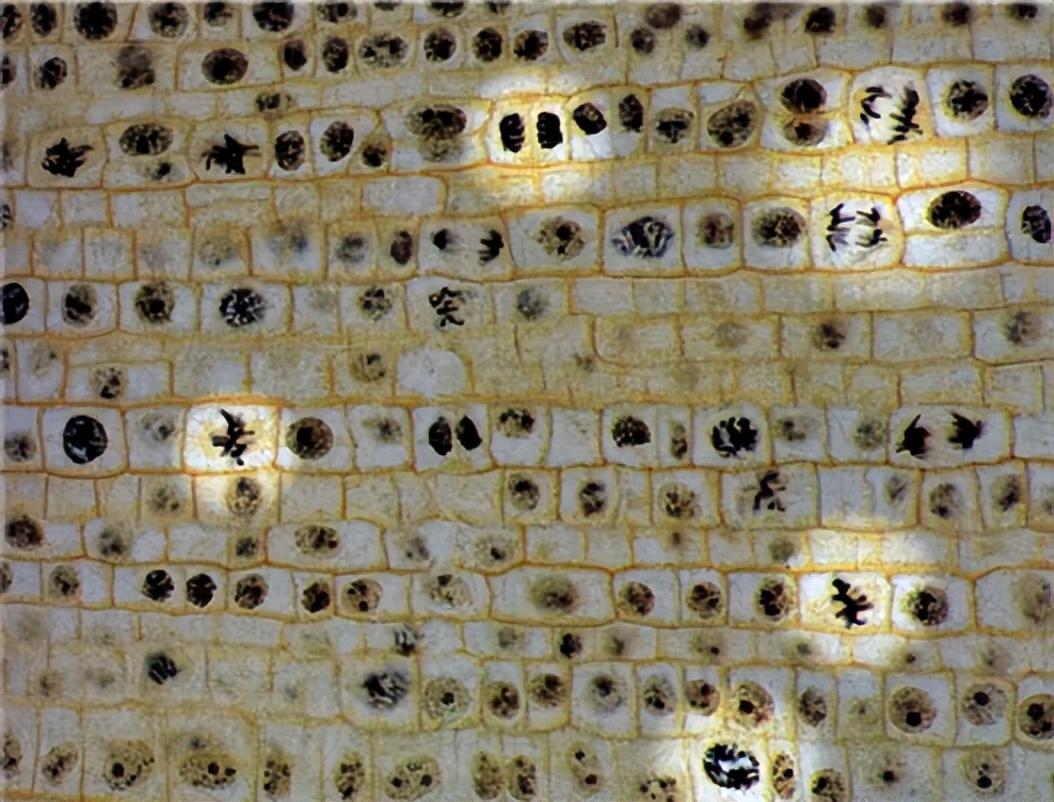
在P2和P3的情况下,正常细胞的比例被计算出来,明显更高(表1)。对P2中遇到的三个异常细胞的临界分析显示,一个细胞发生了腺瘤,另外两个细胞发生了一些结构变化,导致染色体形态明显改变。
相反,P3植株的异常细胞(3)全部记录为非整倍体。在田间转移前分析的P4植株约有97%的正常细胞,而一小部分(3%)的细胞被观察到异常。其中,2%记录为非整倍体,1%记录为结构改变(表1)
在野外条件下生长,对其次生发育的根进行细胞学分析,显示正常细胞数量较多(97.6%)。在2个细胞中表现为腺瘤(2n=31),在1个细胞中表现为多倍体(>4倍)。
另外一个十分重要的发现则表明微核是核不稳定性的主要影响指标因素。微核染色体从未参与数值和/或结构改变,它们在大小和大体形态方面很好地保持了完整性。
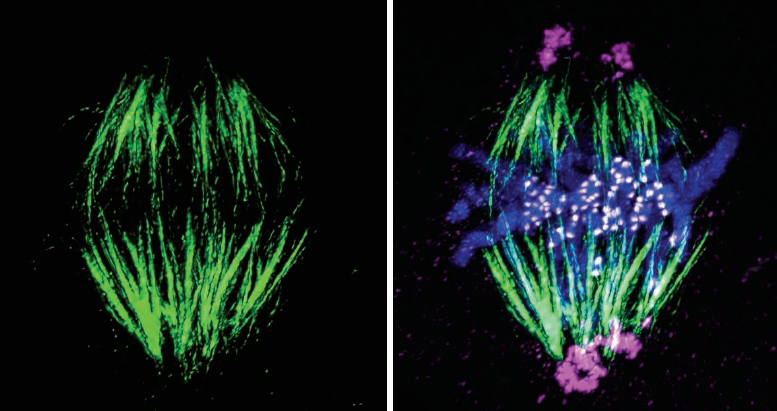
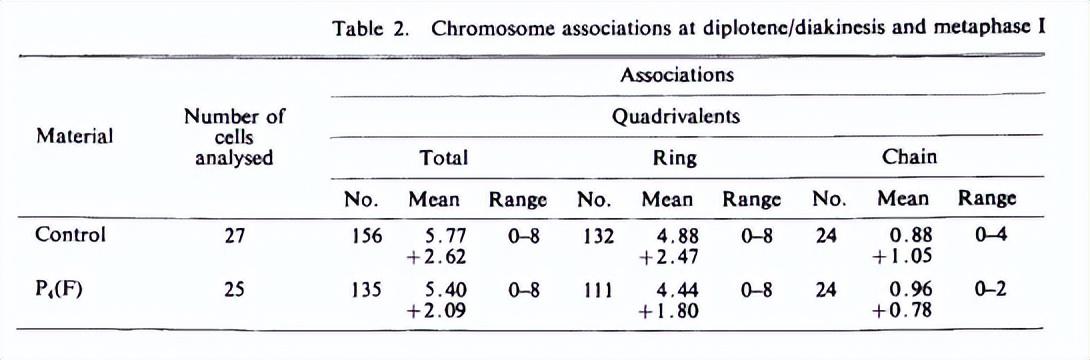
在对照植物中,所有27个花粉母细胞(PMCs)在二倍体或中期I阶段都有四价体、二价体和单价体的混合物,从所有单价体到上述组合(表2,图7)。超过四价体水平的多价体完全不存在。
在pmc中观察到的一个特征是,与二价和单价相比,四价对手的出现比例极高。平均每个细胞有5.77IV 2.6211 3.621(表2)。
每个细胞的四分体数量在0-8之间(表4)。在分析的27个细胞中,多达8个细胞以所有四分体的形式存在关联。
在四分体中,环型占主导地位(平均=4.88;范围=0-8)链条类型(0.88;0-4)(表2)。
这导致一些细胞只有环状四分体,而链型完全不存在。类似地,在二价化合物的情况下,环形结构比棒状(链状)结构更经常遇到。
平均而言,每个细胞有2.18个环和0.44杆二价。虽然在少数细胞中可以观察到多达12个环二价,但记录到的杆二价最多只有两个。
27个细胞中有3个细胞表现出失联,并且在所有3个细胞中都遇到32个单价。
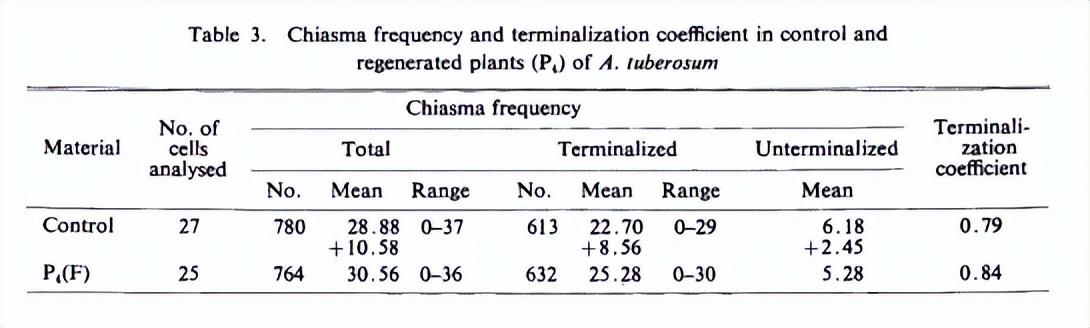
在田间转移P4(F)的再生植株中,双线/终枯期和中期I阶段的染色体关联非常有趣——他们在每个细胞不同关联的平均值上表现出明显的变化。
在分析的25个pmc中,除一份样本有32个单价外,其他所有的pmc都显示出不同比例的四价、二价和单价,平均每个细胞有5.40IV、4.56II和1.4I。
对这份材料的对比发现,也像对照植物一样,在所有分析的细胞中都没有观察到超过四联体水平的关联。
在四价和二价的情况下,环型和链/杆型结合的比例的相对差异遵循与对照植物中观察到的趋势相似(表2)。
范围=0 ~ 8)的链类型(0.96;0 ~ 2)、环二价(3.68;0-9)的发生率高于二价棒(0.8;0.5)。
在P4(F)植物中观察到的另一个有趣的特征是每个细胞的四分位元数在0-8之间(表4),与对照组29.6%的细胞相比,分析的20%的细胞具有所有四分位元。
对照组平均每个PMC的染色体交叉频率为28.88,其中有22.70个被终止,终止系数为0.79(表3),而P,(F)植物的染色体交叉频率和被终止的染色体交叉数量略有增加。
根据计算得出,每个细胞平均交叉数为30.56个,其中25.28个被终止,在块茎连翘对照和再生植株的pmc中,终止系数为0.84(表3)(表2续)
在对照植物中,在后期I观察到的20个细胞中,有13个呈正态分布(16:16)(表5)。其余的细胞呈不均匀分布和/或一系列异常现象,如滞后/滞后的二价体和染色体的延迟分离。
P,(F)与对照的不同之处在于,P,(F)具有更多的(75%)染色体正态分布的细胞,其余25%的细胞呈不均匀分布或具有上述的一些异常(表5)。
两个细胞在后期II显示了编号为2的微核的存在,而一些细胞除了正态分布外,还显示了染色体的滞后和/或晚期分离。
对照植株有75.06%的可染色花粉,而P,(F)植株有73.07%的可染色花粉(表5)。
染色体的不稳定性是体外培养系统及其再生植株的相关现象,许多研究者已经通过根尖的核学研究证明了细胞核的不稳定现象时有发生,而关于减数分裂期间染色体稳定性和配对行为的报道则较少。
我们对有丝分裂和减数分裂两个方面进行了研究,主要目的是监测通过组织培养再生的植物的遗传稳定性,不仅包括培养传代期间,也将观测时长拉长到其野外建立后。

对于一般的各种葱属植物,特别是野韭菜,无论是从染色体数目、大小、形态、配对行为及其分离模式等方面都鲜有报道,因此我们的实验是在这个方向上的第一次尝试。
如前所述,二倍体和四倍体细胞型在自然界是广泛存在的。
在对照植物的根尖细胞中,绝大多数(99.2%)的多倍体互补体(2n=32)的出现证实了本研究中使用的材料是四倍体细胞型。
一个2n=30的异常细胞通常与倍体水平或8作为基本染色体数目的有效性没有任何关系。
减数分裂分析显示29.6%的pmc中有8个四联体,而Mathur和Tandon的学术报道中宣称有16个。
由此可见,在多倍体类群中,这种染色体数目不正常的小比例染色体是自然存在的。

由表1可以清楚地看出,各代(P1、P2、P3、P4)的再生细胞,即使在田间建立后,也不受培养时间延长的影响,保持原有染色体数目(2n=32)。
更有趣的是,畸变的数量和核型只在一小部分细胞中出现。
这些观察数据表明,培养的外植体(茎基)含有大部分均质细胞,这些细胞能够形成不定芽并再生出具有稳定基因型的植株。
在1 ~ 18个月大的野韭菜四倍体细胞型中我们观察到了如非整倍体、三极性和微核等的染色体异常。
与此相反,这种畸变在一个二倍体类群a. cepa的愈伤组织培养物中则很少出现。
基于这种观察结果,我们可以得出结论,外植体材料的倍性水平确实影响染色体异常的范围和频率。
尽管外植体材料是四倍体,但再生体由于没有间接性愈伤组织,在连续几代中保持了染色体数目和结构的完整性,即使有异常,也仅限于非常小比例的细胞。
在多倍体物种,尤其是自四倍体物种中,无论是自然发生还是人工培育,四价平均降低和相应的二价频率增加在连续几代中是很常见的,并且在许多植物组织培养的相关研究材料中都有记载,如油菜、马鞭草、苋属植物、竹节兰和毛豆属植物。
这种现象经常导致“二倍体”,这反过来又帮助植物适应新的物理和生理环境。虽然很难进行一对一的比较,但在P4(F)植物中观察到的趋势与此高度相似。

表2中关于对照和P4(F)植物中四价和二价的平均值的数据显示前者显著下降(从5.77下降到5.40),并且在二价情况下增加了约57.5%(从对照的2.62上升到P4(F)的4.56)。
由此可以清楚地证明,P4(F)植株存在二倍体现象,尽管程度较轻,但和对照组比起来,再生植株中较小比例的单价似乎间接促进了二倍体化过程。
多价和三价的完全缺失进一步证实了这一点,三价和多价是唯一的其他可能的构型。P4(F)植物的交叉频率较高,表明材料的基因组成可能存在更多的重组。然而,更多的染色体交叉被终止,使得终止系数的值更大。
对照植物的相应值分别为28.88和0.79(表2),在P,(F)植物的后期I染色体平均分布细胞百分比的边际增加(16:16)可能与每个细胞的二价平均增加直接相关,与多价构型相比,通常倾向于以一种更有序的方式分离。
这主要是由于中期I二价的构型简单和合适的取向。正常减数分裂的再生植株(P,(F))花粉着色率低,表明后者不是由于任何染色体的因素。
至此我们可以大胆地总结:基因和生理因素的参与也许发挥了至关重要的作用。
笔者通过利用细胞学技术,对野韭菜再生植株在4代培养和田间建立后的遗传稳定性进行了研究。
再生细胞的核学研究表明,目前的研究发现中大部分(>90%)的细胞具有正常的补体和变异仅局限于一小部分细胞,如瘤变、多倍体和结构改变。
再生植株的减数分裂分析表明,它们大体上与对照植株没有差异,所有参数(关联,交叉频率,终端系数和花粉着色性)与对照植株相比没有显著变化。
四价范围和频率显示再生植物的相当大的减少,进一步证实了细胞学技术在确认再生植物遗传稳定性方面的有效性。
郭仰东,《用组培方法繁殖韭菜雄性不育无性系的研究简报》
杨贺,《植物细胞工程应用及发展前景》
马树彬,《中国韭菜雄性不育系的选育与应用》
张宪省,《韭菜可育和不育花药和花粉发育的细胞学观察》
荆绍凌,《细胞工程在玉米种质改良中的应用》
利容千,《韭菜不同品种的核型分析》
 正在请求数据,请稍候!
正在请求数据,请稍候!Runway推出新的 “Watch” 功能 可分享AI生成的视频
本文概要:1.Runway推出新的“Watch”功能,用户可以分享AI生成的视频。2.Runway首席执行官CristóbalValenzuela表示,好莱坞和AI的讨论需要更加深入。3.Valenzuela强调AI生成的视频需要人类的创造力和控制。站长网2023-08-19 15:25:280000Midjourney生成能力惊人 但研究发现它并不理解自己生成的东西
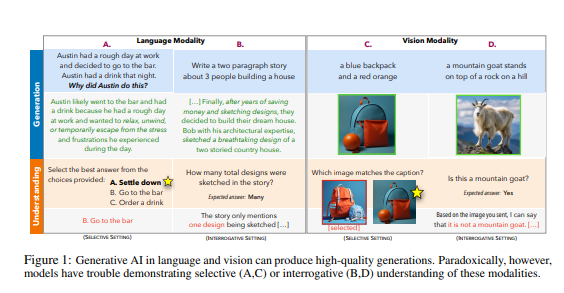
要点:1.生成式AI模型如GPT-4具有强大的生成能力,但在理解生成内容方面存在挑战,引发了一个生成式AI悖论。2.最新研究指出,生成模型的生成能力往往超越了理解能力,与人类的智能配置存在差异。3.这一悖论对人工智能的发展和研究提出了挑战,强调了模型可以创造内容但无法完全理解的概念。站长网2023-11-06 16:59:580000扎克伯格称 Meta 需要数年才能从生成式 AI 赚钱
划重点:⭐️MetaCEOMarkZuckerberg表示,公司需要数年时间才能从生成式AI中获利。⭐️Meta正计划通过广告、订阅等多种方式从其新ChatGPT竞争对手中获利。⭐️MetaAI助手已在Instagram、Facebook和WhatsApp中推出,公司看到多种途径来商业化这一产品。站长网2024-04-26 00:47:390000AI元年,被C端用户们“证伪”了的AI产品(上)
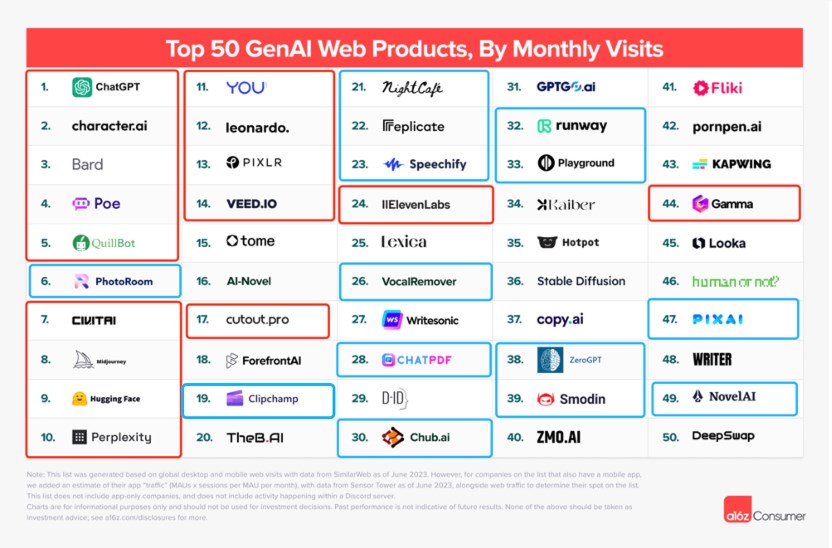
如今半年过去了,市场上AI产品的供给和用户需求又都发生了变化。近日,AI产品榜(aicpb.com)同样按网站流量排出了12月全球Top100AI产品,与a16z上半年榜单相比,变化明显。站长网2024-02-23 14:11:100000AI视野:大模型最快推理芯片Groq登场;真人视频冒充Sora;Stable Diffusion WebUI Forge推出;字节辟谣推出中文版Sora
欢迎来到【AI视野】栏目!这里是你每天探索人工智能世界的指南,每天我们为你呈现AI领域的热点内容,聚焦开发者,助你洞悉技术趋势、了解创新AI产品应用。新鲜AI产品点击了解:https://top.aibase.com/📰🤖📢AI新鲜事大模型最快推理芯片一夜易主Groq每秒可达500tokens【AiBase提要:】⭐️Groq推出的大模型推理芯片性能突出站长网2024-02-20 15:55:290000